- Jan Jansz, in 1811: Jan Kolkman, geb. Oudleusen 1765, woonde
op 't Kolkmans te Oudleusen;
tr. Janna Hendriks, geb. Oudleusen op het erf Velthuijs, dr. v. Hendrik Geerts en Geertje Jansen. - Arend Jansz, in 1811: Arend Kiekebeld, geb. Oudleusen 28 dec. 1766,
overl. Dalfsen juli 1851;
tr. ald. 20 febr. 1813 Annigke Lamberts, geb. Rechteren onder Dalfsen 8 dec. 1782, overl. ald. 4 mei 1844, dochter van Lambert Hendriks en Janna Hendriks. - Hendrik Janssen Uulders 'op Volkerink', geb. Leusen 6 nov. 1768,
woonde later op 't Nijenhuis te Eerde, overl. Bentheim;
tr. 1e Eefze Janssen Nijenhuis;
tr. 2e Aaltje Roelofs (Waaijenk), geb. Lutten (Hardenberg) 14 nov. 1773 en keerde terug naar de boerderij van Jennigje Arends en woonde vanaf dat moment (1773) met zijn gezin 'op Volkerink'. - Jennigje Jans, geb. Oudleusen 1770, overl. ald. 1772.
- Jannigje Jans, geb. Oudleusen 1773, overl. ald. 1774.
- Jan Willem Jansz, geb. Oudleusen 1775.
- Gerrit Jan Jansen, na 1811*: Gerrit Jan Nevenzel, volgt A.
- Dirk Jansen, geb. Oudleusen 1781, in 1811: onbekend;
tr. Hellendoorn 30 juli 1812 Janna Jansen Braakman. - Gerrit Jansen Gortemaker alias Volkerink, volgt B.
- Gerhard Seine Henry Nevenzel. (Zie VIII.)
- Aaltje Martine (Ilse) Nevenzel, geb. 19 mei 1970; tr. Hilvarenbeek 30 juni 2006 Thomas Franciscus Putters, geb. 2 febr. 1976, zn v. Harry J.F. Putters en Paula Balmer; dr. Maria Johanna Pauline Putters geb. 8 juli 2007.
- Karsten Cornelis Gerrit Nevenzel, geb. Hilversum 13 juni 2004
- Wessel Robert Jan Nevenzel, geb. Hilversum 4 apr. 2006
- Hilde Gertrude Karlijn Nevenzel, geb. Hilversum 28 sept. 2008.
- Genetische drift, zeg maar: toevallige mutaties.
- Evolutionaire aanpassing, bijvoorbeeld via huidskleur aan het klimaat. En
- seksuele selectie: lokale voorkeuren voor aantrekkelijkheid vertalen zich op den duur in fysieke variatie.
Al een aantal jaren ben ik zo nu en dan bezig geweest met het opzetten van een familiestamboom. Tijdens
dat onderzoek kwam ik ook materiaal tegen waarvan me werd verteld dat de werkelijkheid anders was dan de
officiële documenten deden voorkomen.
Wanneer zich dan de mogelijkheid voordoet dit te checken middels DNA-onderzoek, dan grijp je dat met
beide handen aan.
Dat DNA-onderzoek is inmiddels (2008) gebeurd. De uitkomsten zijn verbluffend en de gegevens zijn verwerkt in het boek:
|
Genetische genealogie: een zoektocht in ons DNA-archief door
S. Barjesteh van Waalwijk van Doorn - Khosrovani onder auspiciën van
Koninklijk Nederlandsch Genootschap voor Geslacht- en Wapenkunde
Barjesteh van Waalwijk van Doorn & Co's |
Omdat het geen alledaagse kost is om deze verhandeling te begrijpen, zal er een langere aanloop
mijnerzijds moeten volgen om het een en ander te doorwrochten.
Vooraf even de stamboom zoals die, vooral mede door Xander Volkerink, door archiefonderzoek is opgesteld.
tr. circa 1731 Henrikien Henricks. Zij woonden in Oudleusen te Dalfsen, vermoedelijk op 'het Ulderink'. |
|
ged. Dalfsen 1 jun. 1732, woonde te Oudleusen, overl. Dalfsen 13 sept. 1785; tr. Dalfsen 13 mei 1764 (ondertr. ald. 20 april 1764) Jennigje Arends, geb. in de buurtschap Diffelen onder heemse (tegenwoordig Hardenberg) 13 apr. 1738, overl. Beerze 1830, dochter van Arent Gerrits en Wigboldje Geerts; zij hertr. Dalfsen 7 apr. 1786 Willem Janssen en verhuisde naar het erve Volkerink te Beerze, thans Beerzerweg 27. Uit dit huwelijk: |
|
| |
|
Gerrit Jan Nevenzel, tr. 1e Den Ham 22 sept. de weduwe (Willems, Oldensel), tr. 2e Den Ham 19 mei 1830 |
alias Volkerink, tr. Den Ham 6 apr. 1820 |
tr. 27 aug. 1880 |
tr. Dalfsen 2 april 1864 |
|
tr. Den Ham 7 mrt 1889 |
tr. Dalfsen 15 mrt 1900 |
|
tr. Den Ham 14 seot. 1928 |
tr. Dalfsen |
|
tr. Hellendoorn 18 aug. 1967 Uit dit huwelijk: |
|
|
sw. Hilversum 24 juni 2001 Uit dit huwelijk: |
|
- * De Burgerlijke stand is in het grootste deel van Nederland
ingevoerd in 1811 door Napoleon Bonaparte.
Toch bleven er nog geruime tijd mensen over die geen vaste achternaam voerden. Op 17 mei 1813 werd een nieuw decreet uitgevaardigd waarin bepaald werd dat men toch echt voor 1 januari 1814 een achternaam moest kiezen. Maar ook dat werd niet overal opgevolgd.
Veel later vaardigde koning Willem I op 8 november 1825 nog een Koninklijk Besluit uit waarin werd bepaald dat wie nog geen vaste achternaam had vastgelegd, daar nog 6 maanden de tijd voor kreeg.
Hoogleraar Peter de Knijff is samen met o.a. Spencer Wells mede grondlegger van het baanbrekende
Y-chromosomaal onderzoek.
De Knijff is populatiegeneticus aan het Leids Universitair Medisch Centrum (LUMC) en hij deed daar
zijn ontdekking in 1995, min of meer toevallig gelijktijdig met nog anderen.
De simpele aanzet daarvan is: Het afnemen van wat wangslijmvlies. En daar is waarlijk heel wat aan te onderzoeken.
Als je met een wattenstaafje langs de binnenkant van je wang strijkt, sleur je een heleboel wangslijmvliescellen los. Met een beetje rode kleurstof voor wat meer contrast in deze transparante cellen krijg je dan het volgende microscopische beeld (fig. 1) te zien. Wat rommelig, immers de wangslijmvliescellen zijn niet zachtzinnig verkregen. Niet dat dat erg is voor het wangslijmvlies, tijdens bijv. het eten van droge broodkorst krijgt het slijmvlies heel wat meer te verduren. De regeneratie van het vlies is heel sterk.
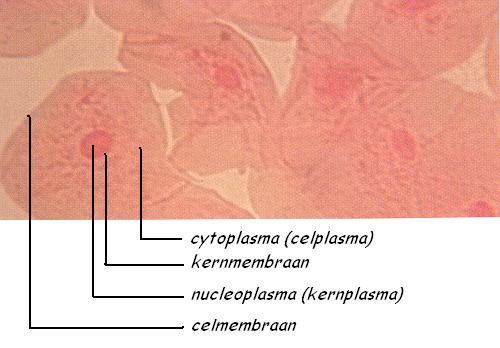
|
In de kern (nucleus) van een wangslijmvliescel zitten 46 kleurbare (chromo) lichaampjes (somata) de chromosomen (kleurlichaampjes), o.a. het Y-chromosoom, althans als het een slijmvlies van de man betrof.
Tijdens een celdeling (cytokinese) moet vooraf een kerndeling (mitose) plaatsvinden en daartoe
moeten de langgerekte en actieve chromosomen in een compacte inactieve transportvorm gebracht worden.
Alleen in deze compacte, ineengedraaide vorm is het mogelijk de chromosomen over de dochtercellen
te verdelen.
Vooral in deze compacte vorm zijn de chromosomen met speciale, aan het DNA hechtende kleurstoffen,
goed als gekleurde lichaampjes zichtbaar te maken.
Officieel moeten het er 46 zijn, mits ze niet teveel over elkaar heenliggen in het preparaat (fig. 2).

|
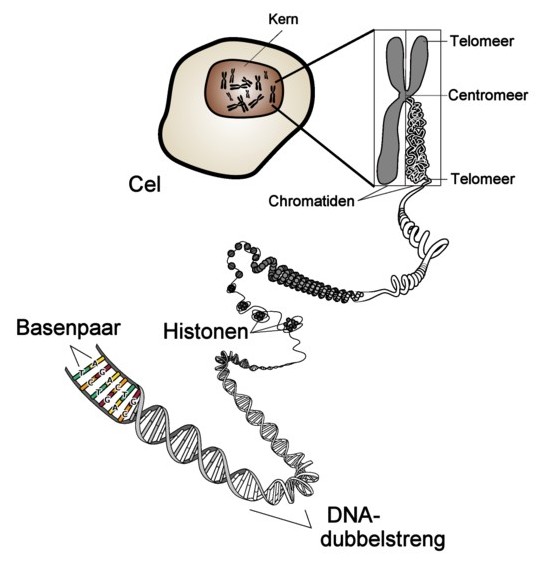
Onder verdere uitvergroting en met weglating van een deel der 'verpakkingseiwitten' (histonen) ziet een (transport-)chromosoom (fig. 3) er schematisch zo uit. Bedenk dat 10 μm (10 mikrometer) één honderdste is van een millimeter.
 |
|
De linkerhelft en rechterhelft noemen we chromatiden(1). Zij zitten nog vast aan elkaar in het middendeel (centromeer)(2). De korte arm en lange arm worden aangeduid met (3) en (4). |
In de celkernen (niet in die van ei- of zaadcellen) komen de chromosomen in paren voor. De 46 chromosomen moeten we dus zien als 23 paar. Bij de vorming van voortplantingscellen worden in een speciale deling (meiose) deze 23 paren netjes gescheiden, zodat in de eicellen en zaadcellen 23 chromosomen voorkomen.
De kernen van lichaamscellen bevatten 23 paren, een dubbel pakketje dat we een diploïd setje noemen;
voortplantingscellen bevatten een enkelvoudig pakketje, een haploid setje chromosomen.
Het laatste paar, de 23-ste chromosomen, bepaalt het geslacht. Bij de vrouw zijn dat twee gelijksoortige en vrij grote chromsomen (gemakshalve XX genoemd), maar bij de man is het ene een sterk gereduceerd chromosoom (gemakshalve XY genoemd). Hieronder staat een elektronenmicroscopische opname (fig. 4) van een X- en een Y-chromosoom, zoals ze bij de man voorkomen.
 |
|
Elektronenmicroscopische opname van het X-chromosoom (links) en het veel kleinere Y-chromosoom (rechts). |
Voor de zekerheid is hieronder nog het chromosomental van 23 voor de man i.c. de zaadcel weergegeven. Een vrouw heeft in de eicel dus 22 + X en en man in de zaadcel 22 + X óf 22 + Y. In de afbeelding hieronder (fig. 5) is dat weergegeven.
 |
|
chromosomen doorgeven, plus óf de X óf de Y. In een zaadcel zit dus 22+X óf 22+Y. Chromosomen worden gerangschikt op lengte en op ligging van het centromeer. |
Het materiaal in de chromosomen dat de erfelijke eigenschappen in zich draagt is het DNZ (desoxy-ribo-nucleïne-zuur. Dit is een heel ongebruikelijke afkorting geworden, iedereen spreekt over DNA, de Engelse afkorting van deoxyribonucleic acid; zelfs de Duitsers raken van hun DNSäure af, ofschoon de Fransen wat hardnekkiger zijn met hun ADN (Acide DésoxyriboNucléique).
|
|
De vaak grillige wangslijmvliescellen bevatten een kern (nucleus), waarin de chromosomen zichtbaar kunnen worden gemaakt. Het DNA in het chromosoom zit verpakt in eiwitten (histonen). Het DNA is een dubbele spiraal (helix) waarin de stikstofbasen A-T en G-C (en v.v. T-A en C-G) tegenover elkaar zitten. (centromeer = middendeel; telomeer = einddeel) |
Een kind erft van beide ouders een enkel pakketje (het haploïde setje van 23 chromosomen). Als
zowel de vader als de moeder het X-chromosoom afgeven wordt hun kind een dochter: XX.
Als de vader het Y-chromosoom doorgeeft dan krijgt het kind een X en een Y. De moeder bezit immers
alleen twee X-chromosomen. De combinatie XY wordt een zoon. De man bepaalt - weliswaar door toeval - welk
geslacht zijn kind heeft.
Al het overige erfelijk materiaal wordt van ouders naar kind in de pakketjes 1 tot en met 22 gekopieerd
en doorgegeven. Bij dit kopiëren wordt het DNA genetisch gezien nogal door elkaar geschud
(er hangt immers geen labeltje aan bijv. het 8-ste chromosoom, óf dat van vader óf moeder afkomstig was),
waardoor het na enkele generaties lastig terug te herleiden is naar een stamvader of -moeder.
Het is als de verdeling van 22 paar blokjes (roze en blauw). Gemiddeld mag je bij verdeling van de paren
verwachten dat de helft aan roze + de helft blauwe in de eene helft (i.c. cel) komt (het meest
waarschijnlijke), maar in de ene helft alleen maar roze en in de andere helft alleen maar blauwe
(het minst waarschijnlijke) is zeker niet uitgesloten.
En dan is er nog iets en dat noemt men recombinatie. Voor de evolutie van onmisbaar
belang!
Bij het in elkaar spiraliseren van dat sliertige chromosoommateriaal om het in de transportvorm
te krijgen, gebeurt het nogal eens dat de chromatiden van een paar verstrengeld raken.
Bij het sorteren en verhuizen breken dan stukken van de chromatiden af.
Dat wordt weliswaar fluks gerepareerd, maar het reparatiemolecuul ziet niet het verschil tussen
een pa- of een ma-DNA-deel. Kortom in de helft van de verstrengelingen zitten die gelijksoortige
stukken verwisseld. Soms (ca. 3 % van de delingen) tref je het en zie je door de microscoop
de chromatide'beentjes' kruiselings over elkaar liggen. Deze overkruising staat bekend onder
de naam crossing over.
Door deze 'fout' kan een toevallig sterke eigenschap van moeder hergegroepeerd (gerecombineerd)
worden met een sterke eigenschap van vader (bijv. 'gevoel voor muziek' gerecombineerd met een 'betere
toetsaanslagmotoriek').
Maar dat geldt niet voor het mannelijke Y-chromosoom. Er is er maar een van. Dus van 'vervuiling' met een ander pakketje tijdens het kopiëren kan geen sprake zijn. Het Y-chromosoom blijft zodoende vele generaties van vader op zoon ongewijzigd, maar ... toch niet helemaal.
Het blijkt namelijk dat er zo heel nu en dan toch spontaan zeer kleine veranderingen, mutaties,
optreden in de chromosomen, dus ook in het mannelijke Y-chromosoom.
Voor het tussenstuk van het Y-chromosoom is dat zo eens in de paar duizend jaar.
Waarom tussenstuk? Dat is het gedeelte dat geen actieve genen bevat. De
uiteinden coderen voor zo'n 27 eiwitten, die vooral verantwoordelijk zijn voor de productie van zaadcellen
en merkwaardigerwijs ook voor de beharing van het oor, op Sri Lanka een markante beharingsvorm,
uiteraard alleen bij mannen.
De mensheid is de afgelopen 200.000 jaar vanuit Afrika over de wereld uitgewaaierd. Dat ging langzaam. Steeds werd een nieuw gebied ontgonnen en steeds duurde het vele generaties eer enkele mensen van een groep weer verder trokken en weer een ander gebied in bezit namen.
Het Y-chromosoom van de mannen in een bevolkingsgroep die vanuit Afrika bij voorbeeld voor het eerst in zuidwest Azië was neergestreken is dus na enkele duizenden jaren veranderd op een klein onderdeel, een genetisch foutje, een mutatie.
Een deel van deze groep vertrekt vervolgens met medeneming van deze mutatie na lange tijd weer,
bijvoorbeeld naar het huidige China, alwaar na een paar duizend jaar bij de mannen in het Y-chromosoom
weer een genetisch foutje optreedt.
Dit foutje kan zich niet meer mengen met de eerste groep, daarvoor leven ze te ver van elkaar verwijderd.
Ze zijn uniek, want ze hebben de oude mutatie + een nieuwe.
En zo valt er - in de mannelijke lijn - een nauwkeurig genetisch spoor over de hele aarde te trekken.
Het Y-chromosomaal-onderzoek heeft dus niet zozeer direct met ras te maken, maar eerder met de regio
waar iemands voorouders hebben gewoond.
Het ras wordt voornamelijk bepaald door de andere 22 paar chromosomen, waarin veel meer DNA zit.
Om die genetische afwijkingen in het Y-chromosoom per regio precies te bepalen heeft onderzoeker De Knijff de afgelopen tien jaar een groot aantal laboratoria over de hele wereld verzocht om DNA-matariaal uit de eigen regio op te sturen.
Hij heeft op die manier inmiddels een databank met 40.000 profielen aangelegd en zo een soort genetische wereldkaart gemaakt.
Voor de families Volkerink en Nevenzel ziet die wereldkaart er aldus uit:

|
Waarom die wereldkaart voor de deelnemers (probandi) Nevenzel en Volkerink er hetzelfde uitziet?
Dat was al eerder via de rijksarchieven uitgezocht. Zij hebben een gemeenschappelijk voorvader in
de persoon van Jan Jansen (Ulderinks), gedoopt te Dalfsen 13 september 1738.
Deze Jan had 7 zoons die na 1811, na de officiele naamsaanneming (definitief in 1825), deels verder
gingen onder een niet meer wisselend patroniem (vaders voornaam), maar onder een vaste (boerderij)naam
i.c. achternaam, te weten: Kolkman, Kiekebeld, Volkerink en
Nevenzel.
Vanaf 1738 tot 2008, een periode van 270 jaar, is er bij beide probandi van Nevenzel en Volkerink in de loop der jaren geen mutatie opgetreden in de bekeken markeringspunten (markers) op het Y-chromosoom en kan met stelligheid beweerd worden dat die voorouders - eufemistisch gezegd - niet vals hebben gespeeld.
Maar wat bedoelen we met die markers 'markeringspunten' op het Y-chromosoom?
Het hele erfelijke DNA-bezit dat we via de 23 chromosomen in de zaadcel meekrijgen noemen we
het genoom. Het genoom van moeder in de eicel en het genoom van vader in de zaadcel levert
na de bevruchting het genotype, de erfelijke aanleg voor het individu dat uit
uit deze kiemcel of zygote zal groeien.
Hoe het individu er later uit zal zien, zijn fenotype hangt af van een groot aantal externe factoren,
zoals een goede innesteling in het baarmoederslijmvlies, goede doorbloeding van de placenta, geen contact met
ziektekiemen, alcohol of nicotine, een voorspoedige bevalling en gunstige factoren voor een goede verdere
ontwikkeling.
Het genoom bestaat uit ca. 3 miljard nucleotiden, de genetische eenheden (of letters) waarin onze te verwerven aanleg wordt weergegeven.
Nucleotiden zijn de DNA bouwstenen en bestaan uit drie onderdelen, nl. een desoxyribose (een suikermolecule met 5 C-atomen), een fosfaatgroep en één van de 4 stikstofbasen die de letter bepalen van het nucleotide.
 |
|
Blauw, rood, groen en paars: De basen G, C, A en T. Oranje: deoxyribosegroep. Geel: fosfaatgroep. De dubbele en driedubbele waterstofbruggen zijn aangegeven met stippellijntjes. De 3'- en de 5'-uiteinden van de "stijlen of bomen" staan eveneens aangegeven |
De stijlen of bomen van de DNA-ladder (DNA-dubbelstreng) worden gevormd door afwisselend een P (phophaatgroep) en D (desoxyribose). De sporten van de DNA-ladder worden gevormd door paren stikstofbasen. Aan hun molecuulstructuur is te zien dat alleen A tegenover T past (2 waterstofbrug-verbindingen, H-bruggen) en G tegenover C (3 H-bruggen). De sporten liggen wat verdraaid t.o.v. elkaar waardoor de ladder in een spiraal of helix wordt gedwongen (Kijk even terug bij fig 6.). Bij het aflezen van een code wordt slechts gebruik gemaakt van één streng, stijl of boom.
Onze erfelijke aanleg wordt maar in vier letters A, T, G, of C neergeschreven.
Hoewel het Y-chromosoom een klein chromosoom is, bevat een DNA-streng toch nog
zo'n 60 miljoen letters, dus de rest van 3 miljard letters ligt in de andere 22 chromosomen.
Op het FLDO (Forensisch Laboratorium voor DNA Onderzoek) zijn de wangslijmvliesuitstrijkjes van
de probandi Nevenzel en Volkerink onderzocht. Dat van Nevenzel kreeg de samplecode 254HH34.
Hieruit werd het Y-chromosomaal profiel vastgesteld.
Dit profiel bestaat uit twee verschillende onderdelen, een haplotype en
een haplogroep.
Een gedeelte uit het middenstuk van het Y-chromosoom wordt voor de profielbepaling gecontroleerd.
Zie het schema hieronder.

|
Het haplotype wordt bepaald door middel van STRs
(Short Tandem
Repeats, 'herhaling van korte gelijksoortige stukken').
STRs zijn dus korte stukjes DNA waarin een specifiek "blokje" van nucleotiden (letters) een aantal
malen achter elkaar (Tandem) herhaald voorkomt (herhalingen - Repeats).
STR is ook de wetenschappelijke Engelstalige naam voor een hypervariabel gebied op het DNA,
bestaande uit deze zich herhalende, repeterende, kleine stukjes DNA.
Een verschil in het aantal herhalingen leidt tot een verschil in de lengte van de STR. De algemene term voor een lengtevariant is allel (mv. allelen. De term is eigenlijk een afkorting van allelomorph 'de andere vorm' die ook mogelijk is van het gen. (Zie verder Genetica)
Voor een STR komen altijd meer dan 2 allelen voor in een populatie. Een populatie is een groep organismen die voor vruchtbare nakomelingen kan zorgen, mits de leden wel in elkaars nabijheid zijn. Ieder allel wordt genoemd naar het aantal voorkomende herhalingen.
Het haplotype bestaat daardoor uiteindelijk uit een reeks getallen, welke overeenkomen met de voor de
verschillende STRs waargenomen allelen.
In het DNA van het Y-chromosoom zijn Segmenten aan te wijzen met het aantal
repeats, bijv. Xander en ik hebben in het segment 19 allebei 16 repeats en dat wordt
genoteerd als DYS19 - 16. Zie fig. 10.
De laatste van de 16 segmenten lijkt een vreemde eend in de bijt, maar is het niet. Het is een tandempje
van de erfelijke letters G-A-T-A en daarvan hebben we 12 blokjes (ook wel satellieten genoemd, kortom
'begeleidende' blokjes. Als de blokjes klein zijn - 3 tot 4 letters - heten ze microsatellieten en
bij 10 tot 60 letters spreken we van minisatellieten). DYS19 is een microsatelliet.

|
In het boek Zonen van Adam staan uitsluitend deze 16 getallen (16, 13, ....., 24, 12) als haplotype bij de probandus (kwartierdrager) onder de haplogroep. Dit kan zo, omdat van deze volgorde niet wordt afgeweken.
De haplogroep wordt bepaald door een serie puntmutaties samen
te nemen. Een puntmutatie is een verandering
van één nucleotide (letter) op één plek (ook locus genoemd). Je kunt het ook anders zeggen, nl.
dat er verschillende
nucleotiden (letters of molecuulvormen) op dat ene plekje kunnen voorkomen en daarom spreken we ook van
SNPs (Single Nucleotide Polymorphisms); van dat enkele (single)
nucleotide komen meer (poly) vormen (morfen) voor.
Een SNP is wel een unieke verandering van één nucleotide naar een ander nucleotide (bijv. als een C verandert
in een T). Ook bij een SNP spreken we van verschillende allelen, maar hier is een allel geen lengtevariant,
maar een verschil in 1 nucleotide.
Een SNP heeft dus minimaal twee allelen en maximaal vier.
De haplogroep bestaat dus uiteindelijk uit een reeks letters, welke overeenkomen met de voor de verschillende SNPs waargenomen allelen.
Volgens wereldwijd aangenomen afspraken worden deze letterreeksen vervangen door een eenvoudiger naamgeving, bestaande uit een aantal hoofd-haplogroepen (A t/mT), welke verder kunnen worden onderverdeeld in sub-haplogroepen, bijv. J1 en J2.

|
In figuur 9 wordt het verschil tussen een STR en een SNP schematisch weergegeven. In deze figuur is één
stukje van het Y-chromosoom van 5 verschillende mannen weergegeven.
In dit stukje Y-chromosoom bevinden zich 1 STR (met de repeat CTAG) en 3 SNPs (een C/T SNP
links van de STR en een A/C SNP en een A/G SNP rechts van de STR).
|
|
Personen kunnen één of meerdere typeringen delen.
Zo delen mannen 1, 4 en 5 het allel 4 (d.w.z. 4 blokjes) voor de STR (haplotypering).
De mannen 1, 2 en 3 het allel C voor de eerste SNP (uit de haplogroepering).
Echter de combinatie van haplotype (STR) en haplogroep (SNPs) is in ieder van deze 5 mannen
uniek, waardoor kan worden geconcludeerd dat zij niet in de directe mannelijke lijn
aan elkaar verwant kunnen zijn.
Voor sample 254HH34 van Nevenzel is, op basis van de typering van 24 SNPs, de haplogroep J waargenomen. Deze is identiek voor Volkerink. Voor de migratieroute - zie beneden - is dat de marker M89. Voor de haplogroep J2 is dat de marker M172.
Figuur 11a geeft een globaal overzicht van de wereldwijde verspreiding van deze haplogroep: Hoe donkerder een regio gekleurd is, hoe vaker haplogroep J in deze regio wordt aangetroffen. Hiermee wordt eveneens de kans groter dat de donor van sample 254HH34 voorouders in de directe mannelijke lijn heeft die uit zo'n 'donker gekleurde' regio afkomstig zijn.
Figuur 11b laat de verspreiding van dezelfde haplogroep binnen Europa zien. Beide kaartjes zijn gebaseerd op een nog niet publiek toegankelijke database van > 60.000 wereldwijd verzamelde mannen. Deze database is eigendom van het FLDO en niet toegankelijk voor "derden".
Uit een verdere subtypering van sample 254HH34 is gebleken dat dit sample tot de subhaplogroep J2 behoort. Echter, op basis van de huidige databestandgegevens is het nog niet mogelijk een gedetailleerde kaart te maken van de verspreiding van deze subhaplogroep. Derhalve is er voor gekozen de verspreiding van de gehele haplogroep J weer te geven.
Het vertrek van haplogroep J (M89) gebeurde zo'n 45.000 jaar geleden vanuit Afrika naar het
Midden-Oosten (Zie migratiekaartje hieronder). Uit haplogroep J onstond zo'n 10.000 geleden
de haplogroep J2 (M172) die een uitloper naar Italië heeft.
Nu blijkt dat er in N.O.-Brabant een cluster J2 zit, zo tegen de Maas aan.
Verder is er een cluster J2 te vinden bij de muur van Hadrianus in Engeland.
Een snelle conclusie is dan dat deze J2-voorouders van Nevenzel en Volkerink afstammen van
(huur)soldaten uit het Romeinse leger, die aan de limes waren gelegerd.
Dat een voorvader vanuit Brabant naar Dalfsen (Ov.) is uitgezwermd is zeker niet uitgesloten.
De limes (Lat. grens) is de aanduiding van de grens van het Romeinse Rijk.
Deze liep van de Noordzee langs de Rijn en Donau naar de Zwarte Zee.
Ofschoon het woord limes strikt genomen zou moeten slaan op de eigenlijke grens aan de Rijn,
hadden de Romeinen ook verdediging in de diepte, zodat het bestaan van versterkingen aan de Waal
en Maas aannemelijk is. Verschillende stenen resten in de Maas zijn wel als militair geduid.
Bij Cuijk zijn de resten van een tijdelijk kamp gevonden.
In de Late Oudheid werd deze diepteverdediging uitgebreid met versterkte bruggen in bijvoorbeeld
Maastricht en opnieuw Cuijk.
Overigens moet bedacht worden dat nagenoeg ieder castrum (vesting, militaire post) en
elk castellum (fort, blokhuis) omringd werd door een dorp (vicus) dat dienstbaar
was aan het fort (waaronder kinderen van Romeinse vaders die landbouw bedreven).
Nog iets anecdotisch. Er hoeft totaal geen relatie te zijn met de J2-groep.
222 - 235 is de regeringsperiode van de Romeinse keizer Alexander Severus.
Uit deze periode zijn twee steles (grafmonumenten) opgegraven (17 nov. 1863) te
Vercovicium (nu Housestead) bij de Muur van Hadrianus. De tekst op stele:
1. Deo Marti et duabus Alaisiagis et N(umini) Aug(usti) Ger(mani) cives Tuihanti cunei Frisiorum Ver(covicianorum) Se(ve)r(iani) Alexandriani votum solverunt libent(es) m(erito).
Vertaling
Aan de god Mars en de twee Alaisiagae en de goddelijkheid van de keizer hebben de Germanen uit de civitas Twente, in de Cuneus Severianus Alexandrianus der Friezen te Vercovicium, hun gelofte ingelost, gaarne en met reden.
 |
